Key words
Antinutritional factors, plant raw material, fish
Giri?
Karma yemlerin bal???n bütün ihtiyaçlar?n? kar??layacak ?ekilde düzenlenmi? olmas?, bal???n sa?l?kl? ve h?zl? büyümesindeki ba?ar?y? art?ran bir unsurdur. Bu nedenle karma yem formülüne girecek yem ham maddelerinin seçimi büyük önem ta??maktad?r (Aky?ld?z, 1992).
Bal?k unu yüksek düzeyde protein içermesi, dengeli bir aminoasit kompozisyonuna sahip olmas? ve bal?k taraf?ndan lezzetli bulunmas? nedeniyle bal?k yemlerinde kullan?lan vazgeçilmez bir protein kayna??d?r. Artan talep ve kümes hayvanlar? ile büyükba? gibi çiftlik hayvanlar?n?n yemlerinde de kullan?lmas?ndan do?an rekabet ile dünya bal?k unu üretimindeki aç?k ikiye katlanmakta ve fiyatlar? daha da art?rmaktad?r. Bunun sonucunda da bal?k ununun artan fiyat? ve elde edilmesindeki belirsizlik, bal?k unu yerine geçebilecek daha ucuz ve kolay elde edilebilen bitkisel protein kaynaklar?n?n, bal?k yemlerinde kullan?mlar?n? zorunlu hale getirmektedir. Ancak bitkisel hammaddeler çe?itli antibesleyici yani beslenmeyi s?n?rland?r?c? faktörler içermektedirler. Bu faktörler sayesinde bitkiler, do?ada kendilerini fare, böcek, bakteri ve ku? gibi canl?lar?n sald?r?lar?na kar?? korumaktad?rlar (Kaya ve Yalç?n, 1999).
Antibesleyici faktörlerin çe?iti ve etkisi, bitkisel hammaddenin cins, tür, varyete ve bitkinin de?i?ik k?s?mlar? ile geli?me dönemlerine göre farkl?l?k göstermektedir. Ayr?ca hayvan türüne göre de antibesleyici faktörlerin etkileri farkl? olmaktad?r (Balabanl? ve ark., 2006). Bu faktörler bal?klarda olumsuz etkiler yapmas?na ra?men, besinsel de?erini art?rmak için bir tak?m i?lemlerden geçirildi?inde büyük bir k?sm? tahrip olmakta veya inaktif hale geçmektedir.
Bitkisel hammaddelerin içerdi?i bu faktörler ve etkileri dikkate al?nd???nda, özellikle entansif bal?k yeti?tiricili?inde kullan?lan yemler ve kullan?lan hammaddeler konusunun önemi ön plana ç?kmaktad?r. Çünkü yeti?tiricicilik i?letmelerinde, i?letme giderlerinin %50 sini veya daha fazlas?n? yem giderleri olu?turmaktad?r. Bu nedenle ekonomik bir üretim, ucuz ancak besin içeri?i dengeli yemlerin elde edilebilmesine ba?l?d?r. Bu derleme, bitkisel hammaddelerde var olan antibesleyici faktörler ve bu faktörlerin bal?klar üzerindeki etkilerini içermektedir.
Antibesleyici Faktörler
Antibesleyici faktörler ayn? zamanda, antibesinleri yok etmek için kullan?lan en yayg?n yöntem olan termal i?lemlere gösterdikleri dayan?kl?l??a göre; ?s? karars?zlar? (proteaz inhibitörleri, fitatlar, lektinler, goitrojenler ve antivitaminler gibi faktörler) ve ?s? kararl?lar? (saponinler, ni?astas?z polisakkaritler, antijenik proteinler, östrojenler ve fenolik bile?ikler gibi faktörler) ?eklinde de s?n?fland?r?labilirler (Francis ve ark., 2001).
Proteaz ?nhibitörleri
Proteaz inhibitörleri; çözünebilen proteinler (albumin) olup soya, bakla, m?s?r, pirinç, arpa, yerf?st???, patates gibi çe?itli bitkisel dokularda bulunurlar. Bu inhibitörlerin etkileri, kökenlerine ve hedef enzimlerine ba?l?d?r. Bitki orijinli proteaz inhibitörleri üzerinde en çok çal???lan? soya fasülyesindeki tripsin inhibitörleridir. Tripsin inhibitörleri, tohumun kotiledon (çim yapraklar?) k?sm?ndaki proteinde do?al olarak bulunan proteaz inhibitörleridir. Bu proteaz inhibitörleri; endojen proteazlar? engelleyerek depolama esnas?nda proteolitik olaylar?n durmas?na neden olmaktad?rlar (Pek?en ve Art?k, 2005).
Proteaz inhibitörleri, ham yem maddesi veya tam olarak i?lenmemi? baklagil tohumlar?yla birlikte al?nd?klar? zaman sindirim sisteminden salg?lanan tripsin ile birle?erek inaktif formda kompleks bir bile?ik meydana getirirler. Olu?an bu kompleksler ince ba??rsaktaki tripsin, kemotripsin ve amilaz enzimlerinin aktivitelerini bask?layarak, yem proteinlerinin parçalanmas?n? dolay?s?yla aminoasit emilimini ve protein yararlanabilirli?ini azaltmaktad?rlar (Tacon, 1997).
Hemaglutininler (Lektinler)
Lektinler, glikoprotein ve karbonhidratlar? ba?lama yetene?ine sahip protein yap?s?nda bile?iklerdir (Seyrek ve Bildik, 2001). Özellikle baklagillerde ve baz? ya?l? tohumlarda bulunurlar. Ba?l?ca antibesleyici etkisi ince ba??rsakta son ürünlerin emilimini engellemek ?eklindedir. Lektinler, ince ba??rsak epiteli üzerinde bulunan mikrovilli yüzeyindeki glikoproteinlere ba?lanarak baz? yaralar?n olu?mas?na, anormal mikrovilli geli?imine ve buna ba?l? olarak goblet hücre yo?unlu?unda art??a yol açmaktad?rlar (Hunt ve ark., 2007). Makromolekülleri parçalayan ve hücre içine ta??yan enzimleri içeren mikrovillilerin yap?s?n?n bozulmas? ve dolay?s?yla mukus salg?s?n?n artmas? da ba??rsak epitel hücrelerinin enzim ve emme kapasitelerini dü?ürmektedir. Lektinlerin, ba??rsak epitel dokusundaki yap?ya verdikleri zarar sonucunda glikoz, aminoasitler, vitamin B12 ve iyon absorbsiyonu önemli düzeyde engellenmektedir.
Lektinler, nemli ?s? i?lemi (100 0C 10 dakika) ya da otoklavlama i?lemiyle ayr?labilirler (Aregheore ve ark., 1998).
Tanenler
Tanenler genellikle bitkilerin kök, odun, kabuk, yaprak ve meyvelerinde yer alan polifenolik bile?iklerdir. Kolza, bakla, çay ve sorgum gibi bitkilerde yo?un olarak bulunurlar. Tanenler kimyasal aç?dan, hidroliz olabilen tanenler ve kondanse tanenler olmak üzere iki ana grupta incelenirler. Birinci grupta yer alan tanenler bir asit ya da enzim e?li?inde hidroliz olarak gallik asit, pirokate?ik asit ve ?eker gibi suda çözünebilen bile?iklere dönü?ürler. Hidroliz olabilen tanenlerin en iyi bilinen örneklerinden biri gallotanenlerdir. Çok daha geni? bir grup olan kondanse tanenler ise hidroliz olamazlar. Bunlar ?s? kar??s?nda, kuvvetli asitlerle ya da baz? yükseltgeyici maddelerle flovaben denen koyu k?rm?z? renkli çözünmez bile?ikler olu?tururlar (Anonim, 2008).
Tanenler; yem ham maddelerindeki esansiyel mineraller, proteinler ve karbonhidratlarla kompleks bile?ikler meydana getirerek yemin besleme de?erini dü?ürürler ve büyümede gerilemeye neden olurlar (Balaban ve Ark., 2006). Tanenler, substratlara ba?land?klar? ve kompleks bile?ikler olu?turduklar? için tripsin ve amilaz enzimlerinin aktivitelerini olumsuz yönde etkileyerek protein ve ni?asta sindirimini engellerler. Kolza veya kanola küspelerinde bulunan tanenler, küspelerdeki demir ile kuvvetli bir demirfenol kompleksi olu?turarak demirin emilimini önemli derecede azalt?rlar. Ayr?ca B12 vitamini ile de kompleks bile?ikler olu?turarak bu vitaminin emilimine engel olurlar (Hunt ve ark., 2007).
Yap?lan çal??malarda, bal?k türlerinin tanenlere kar?? toleranslar?n?n farkl? oldu?u tespit edilmi?tir. Bu durumun, tanenlerin yap?s?ndaki farklardan ya da tanenlerin yemdeki di?er antibesleyici faktörlerle aralar?ndaki etkile?imden kaynakland??? dü?ünülmektedir. Fish ve Thompson (1991), tanen ve lektin aras?ndaki etkile?imin tanenin amilaz üzerindeki yok edici etkisini ortadan kald?rd???n? bildirmi?tir.
Tanen içeren hammaddelerin, okside edici ajanlarla i?lenmesi ya da polietilen glikol gibi tanen kompleksleyicilerle desteklenmesi, tanenlerin hayvanlar üzerindeki negatif etkilerini azaltmaktad?r (Makkar ve Becker, 1996).
Gosipol
Gosipol; Gossypium (pamuk) cinsine ait bitkilerin pigment bezlerinde ve di?er baz? iki çenekli bitkilerde bulunan, aromatik aldehitlerden olan, ya?da ve alkolde çözünen, koyu renkli polifenolik yap?da ve serbest formu zehirli olan bir pigmenttir. Serbest gosipol toksik oldu?u halde, ba?l? formda olan? toksik de?ildir. Pamuk tohumunun ?s?l i?leme tabi tutulmas? gosipolün ba?l? forma geçirilmesi içindir. Bu esnada gosipolün form grubu ile lisin ve argininin serbest amino grubu veya sisteinin tiyol grubu etkile?erek gosipolü ba?lamaktad?r. Pamuk tohumunun protein olmayan bile?enleri de ?s?l i?lemle serbest gosipolü ba?l? hale getirebilir. Böylece serbest gosipolün düzeyi % 0.04 alt?na dü?mekte ve olu?an konjüge bile?ikler devam eden ?s? nedeniyle moleküler farkl?la?maya ba?lamakta ve sonunda çözünmez, sindirilemez, polimerize ürünlere dönü?mektedirler. Bu ba?lanma olay? gosipolün toksisitesini giderirken sindirilmesi mümkün olmayan gosipol-protein komplekslerinin olu?mas?na, ya? metabolizmas? için büyük önem ta??yan metiyonin gibi baz? amino asitlerin bozulmas?na ve ayr?ca lisin ve arginin gibi amino asitlerin yaray??l?l???n?n azalmas?na neden olmaktad?r.
Gosipol, bal?klarda büyümede gerilemeye, barsak ve di?er iç organ bozukluklar?na (karaci?erde lezyon, böbreklerde hasar) sebep olabilmektedir. Ayr?ca sperm aktivitesinde de?i?ikliklere, anormal spermatozoidlerin olu?umuna, testis histolojisinde anormalliklere neden olarak da üreme sistemini etkilemektedir (Francis ve ark., 2001).
Fitik Asit – Fitatlar
Fitik asit, miyoinositol halkas? ve buna ba?l? inorganik fosfattan olu?an serbest bir ester asididir. Kimyasal ad?, miyoinositol 1, 2, 3, 4, 5, 6 hekzakis dihidrojen fosfatt?r. Fitik asit ve fitatlar, bitki tohumlar?nda, dane yemlerde, kök ve yumrularda yayg?n olarak bulunurlar (Ergün ve ark., 2002). Fitatlar, özellikle de tah?llarda d?? endospermde yo?unla?m??lard?r. Hububatta fitik asidin birikim bölgesi alöron tabakas?d?r (Alöron tabakas?, tanede endospermin en d???ndaki iri ve tek s?ral? hücrelerden meydana gelen kül ve proteince zengin olan, ö?ütme s?ras?nda kepe?e dahil edilerek undan ayr?lmaya çal???lan tabakad?r). Bu?day ve pirinç tanelerinin endospermi hemen hemen fitik asitten yoksundur. Fitik asit bu tanelerin kepek ve ru?eym (tanenin filizlendi?i bölge) tabakalar?nda yo?unla?m??t?r. M?s?rda ise fitik asidin %88’i ru?eymde bulunmaktad?r (Bilgiçli, 2002).
Fitik asit beslenme aç?s?ndan öneme sahip minerallere ba?lanarak (Ca, Zn, Fe, Mn vb.) ince barsakta sindirim ve emilimi daha az olan kompleks bile?ikleri yani fitat bile?iklerini olu?tururarak, minerallerin biyo-yaray??l?l???n? dü?ürmektedir (Richardson ve ark., 1985). Fitik asitin minerallerle kompleks olu?turmas?yla meydana gelen fitatlar, tripsin ve kemotripsin gibi endojen proteazlara ba?lanarak proteolitik enzimler taraf?ndan daha zor parçalanan fitatprotein komplekslerini olu?turmalar? sonucunda protein ve amino asitlerin sindirilebilirli?ini azalt?rlar (Midilli ve ark., 2003). Bu nedenle fitat içeren yemlerle beslenen bal?klarda dü?ük büyüme performans? elde edilmektedir.
Fitin fosforunun de?erlendirilebilmesi için fitik asit molekülünün hidrolize olmas? gerekmektedir. Fitin fosforunun hidrolizi; ?slatma, çimlendirme, bitkisel endojen fitaz enzimince zengin g?dalar? kullanma, depolama, pi?irme ve otoklavlama gibi yöntemlerle gerçekle?mektedir (Pek?en ve Art?k, 2005). Ayr?ca, d?? kabu?un soyulmas? için ö?ütme i?lemi yap?lmas? ve fermantasyon i?lemiyle maya ve laktik asit bakterileri taraf?ndan olu?turulan fitaz enzimi de yemdeki fitat miktar?n? dü?ürmektedir. (Midilli ve ark., 2003).
Glikosinolatlar
Glikosinolatlar yada tioglikosidler, turpgiller ve özellikle kolza tohumu küspesi, hardal tohumu gibi Brassica cinsi üyelerinde do?al olarak bulunan glikozidlerdir. Glikosinolatlar toksik olmad??? halde enzimatik hidrolizleri sonucunda goitrin, izotiyosiyanat, tiyosiyanat iyonlar? ve nitril içeren büyüme inhibitörleri ile goitrojenler gibi toksik ürünler aç??a ç?kmaktad?r (Tacon, 1997). Bu toksik ürünler ise organizman?n iyot al?m?n? dü?ürdükleri gibi tiroit bezinin yap?s? ve fonksiyonunu bozmakta ayr?ca karaci?erde de hasara neden olmaktad?rlar (Ergün ve ar., 2004).
Bal?klar?n glikosinolat içeren yemlerle uzun süre beslenmesi durumunda, bal?klar?n tiroit fonksiyonlar?ndaki bozulmaya ba?l? olarak metabolizmalar? ve büyümeleri de olumsuz yönde etkilenmektedir (Hossain ve Jauncey, 1989). Uygun bir ?ekilde ?s?l i?lem uygulanmas? glikosinolatlar?n toksik etkisini minimum düzeye dü?ürmektedir. Kolzan?n ?s?l i?leme tabi tutulmas?yla mirosinaz inaktif hale getirilerek glikosinolatlar?n toksik ürünlerine ayr?lmas? önlenir (Burel ve ark., 2000).
Antivitaminler
Bal?k yemlerinde kullan?lan soya fasulyesi, yonca ve ya?l? tohumlar?n unu gibi pek çok alternatif protein kayna??n?n, besin kayna?? olarak etkinliklerini azaltacak çe?itli anti-vitaminler içerdi?i bilinmektedir. Antivitamin A, soya fasülyesinde bulunur ve karotenlerin y?k?m?na sebep olur. Antivitamin D, soya fasülyesinde bulunur, civcivlerde kalsiyum ve fosfor absorbsiyonunun engellenmesine yol açar. Antivitamin E, soya fasülyesi, bezelye, börülce ve adi yoncada bulunur, civciv ve kuzularda karaci?erde nekroza ve kaslarda dejenerasyona sebep olur. Antivitamin K, tatl? yoncada bulunur. Anti-tiamin, pamuk tohumu, hardal tohumu, mung fasülyesi ve keten tohumunda bulunur. Anti-niasin sorgumda, Anti- piridoksin keten tohumunda, Antivitamin B12 ise çi? soyada bulunur (Tacon, 1997). Vitaminler ?s?ya duyarl? olduklar?ndan, düzgün ?s? i?leminden geçirilmi? bal?k yemleri kullan?ld???nda, antivitaminlerin bal?klardaki fizyolojik etkilerinin pek önemli olmad??? söylenmektedir (Francis ve ark., 2001).
Alkaloitler
Alkaloitler bitkilerde en fazla yer alan toksik bile?iklerdir ve suda az, organik çözücülerde ise daha fazla çözünürler. Canl? metabolizmas? üzerinde fizyolojik etkilerde bulunan alkaloitler, genellikle karma??k kimyasal yap?s? olan, halka formunda ve azot içeren bitkisel bazlard?r. Bitkilerde hücre öz suyunda erimi? olarak yer alan alkaloitler, genellikle belli bir organda (kök, kabuk, yaprak, meyve, tohum gibi) daha yüksek oranda bulunurlar. Alkaloitlerin ço?u bir türe veya yak?n türlere özeldir, bir k?sm? ise bir familyaya hast?r. Bu nedenle bitkilerde nadiren bir tek alkaloit vard?r. Genelde çok küçük farklarla ayn? yap?ya sahip bir grup alkaloit birlikte bulunur. Bunlardan biri di?erlerinden daha fazlad?r ya da daha aktiftir. Morfin, nikotin, kinin, atropin, kokain ve solanin alkaloitlerden baz?lar?d?r. Alkaloitler sinir sistemi ve karaci?er üzerine etkilidirler. (Ergün ve ark., 2002). Antibesleyici özelli?i sahip alkaloitleri içeren yemlerle beslenen bal?klarda yem al?m?nda azalmalar görülmektedir (De la Higuera ve ark., 1988).
Alkaloitlerin bal?klarda ne çe?it fizyolojik reaksiyon yapt??? henüz tam olarak anla??lamam??t?r ve bu konuda daha fazla çal??maya ihtiyaç duyulmaktad?r.
Saponinler
Saponinler, baklagiller gibi bitkisel kaynakl? yem hammaddelerinde bulunan steroit ya da triterpenoit glikozitlerdir. Genelde ac? tatlar?, sulu çözeltilerde köpürmeleri ve k?rm?z? kan hücrelerini hemolize u?ratabilmeleri ile tan?n?rlar. Küçük kabarc?klar olu?turarak suyun yüzey geriliminin azalmas?na ve atmosferle su kütlesi aras?nda difüzyonun engellenmesine neden olan saponinler, bal?klar taraf?ndan a??z yoluyla al?nd???nda, bal?klar?n sindirim sisteminde parçalanarak sisteme zarar verirler. Dola??m sistemiyle vücuda yay?lmalar?yla da toksik etki gösterirler. Ayr?ca, bu madde suyla kar??t?r?ld???nda deterjan etkisi nedeniyle bal?klar?n solungaç epitellerinde hasara yol açmaktad?r (Erta?, 2007). Saponinleri bitkisel hammaddelerden uzakla?t?rmak için en uygun yöntem suyla ay?rma yöntemidir.
1 gkg seviyesinin alt?ndaki saponin miktar? yeti?tiricili?i yap?lan bal?klar?n büyüme performans?n? genellikle etkilememektedir. Yayg?n olarak bulunan bu maddenin etkileri ve tolerans limitleriyle ilgili daha çok çal??ma yap?lmas? gerekmektedir (Francis ve ark., 2001).
Fitoöstrojenler
Fitoöstrojenler bu?daygiller ve baklagiller grubuna ait birçok yem bitkisinde bulunan, östrojen gibi davran?? gösteren steroidal yap?da olmayan bile?iklerdir. Fitoöstrojenler, yap?lar?na ba?l? olarak hem östrojen hem de östrojen etkisinin tersi etki gösterme potansiyeline sahiptir. Bitkilerde fenilpropan ve basit fenollerden sentezlenen fitoöstrojenler, kimyasal olarak çok geni? çe?itlilik göstermektedirler. Bilinen en iyi fitoöstrojen kaynaklar? Leguminosae ailesine ait bitkilerden kurubaklagiller (bezelye, fasulye, mercimek vb.), özellikle de soya fasulyesidir (Büyüktuncer ve Ba?aran, 2005).
Östrojen, hayvanlar?n çe?itli fizyolojik i?lemlerinde çok geni? etkilere sahiptir, dolay?s?yla yemde kullan?lan bitkisel hammaddelerin östrojenik etkilerinin çok çe?itli sonuçlar? olabilir. Östrojenik etkiye sahip bu bile?iklerin bal?klarda nas?l bir etki yapaca?? konusunda yap?lan çal??malarda, yemde bulunan fitoöstrojenlerin bal?kta östrojenik etkiye sebep olduklar?, vitellogenesise yol açt?klar? ve dü?ük büyüme oran?na sebep olduklar? görülmü?tür (Kaushik ve ark., 1995; Pelissero ve ark., 1991). Fitoöstrojenlerin gözlenen etkilerinin daha iyi de?erlendirilebilmesi için bal?klarda daha fazla çal??ma yap?lmal?d?r.
Oligosakkaritler ve Ni?astas?z Polisakkaritler
Oligosakkaritler ve ni?astas?z polisakkaritler baklagil ve tah?l tanelerinde bulunurlar. Bal?klardaki negatif etkileri ya safra asidine ba?lanma yada sindirim enzimlerine kar?? yap?lan hareketi k?s?tlama ve ba??rsaklarda substratlar?n hareketini t?kama ?eklinde ortaya ç?kmaktad?r (Storebakken ve ark., 1998).
Baklagil tohumlar?na yüksek s?cakl?klarda uygulanan ekstrüzyon yöntemi, hücre duvarlar?n?n daha fazla parçalanmas?na veveya alfa galaktozitlerinin k?smen parçalanmas?na (ayr??t?r?lmas?) sebep oldu?undan dolay? karbonhidrat sindirilebilirliklerini artt?rmaktad?r. Ni?astas?z polisakkaritlerin, özellikle de çözünebilir olanlar?n bal?klar?n büyümeleri için oligosakkaritlere oranla daha zararl? olduklar? söylenebilir (Refstie ve ark., 1999). Ni?astas?z polisakkaritlerin beslenmeyi s?n?rland?r?c? etkileri, suyu hapsederek ba??rsaklarda sak?z?ms? kütleler olu?turmalar?yla ba??rsak içeriklerinin yap??kanl?klar?n? artt?rarak sindirim enzimlerinin aktivitesini k?s?tlamalar?ndan kaynaklanmaktad?r.
Antijenik ?çerikler
Baz? tah?l ve baklagillerdeki protein içeriklerinin hayvanlarda antijenik etkiye sebep olduklar? bilinmektedir. Krogdahl ve ark., (2000) solventle yada alkolle ekstrakte edilmi? soya fasülyesi içeren yemlerle beslenen Atlantik salmonlar?n?n ba??rsak mukozas?nda lezyonlara, villilerde anormalliklere, spesifik ve spesifik olmayan ba????kl?k sistemi tepkilerine sebep oldu?unu bildirmi?lerdir. Gökku?a?? alabal?klar? üzerine yap?lan bir ara?t?rmada ise soya içerisinde bulunan antijenik içeriklerin bal?klar?n büyümesini negatif yönde etkiledi?i saptanm??t?r (Rumsey ve ark., 1993).
Di?er Antibesleyici Maddeler
Siyanojik Maddeler: Bal?k yemlerinde hammadde olarak kullan?lan taneli baklagillerde, kasava gibi kök çe?itlerinde ve keten tohumu gibi ya?l? tohumlarda yüksek miktarda bulunurlar. Siyanojenler, hidrolize edildiklerinde hidrojen siyanür ve di?er karbonil bile?ikler gibi, do?al solunumu engelleyen ve kalp krizine yol açan, zehirli ürünler üretmektedirler. Bu üretim için gerekli olan enzimler genellikle bitki dokular?nda hücre zar? d???nda bulunurlar. Keten tohumu ve kasava gibi siyanojen içeren yemlerle beslenen bal?klarda dü?ük büyüme performans? gözlenmi?tir (Hossain ve Jauncey, 1989).
Mimosin: Leucaena leucocephala bitkisinde bulunan nadir bir amino asittir ve toplam proteinin kuru a??rl???n?n % 3-5’ini olu?turur. Bal?klarda, dü?ük büyüme ve yem de?erlendirmenin yan? s?ra üreme sisteminin bozulmas? ve teratojenik etkiler gibi pek çok olumsuz etkiye sahiptir. Yap?lan ara?t?rmalarda yemdeki Leuceana yapra?? proteinlerinin bal?klar taraf?ndan iyi tolere edilemedi?i ve erkek tilapya bal?klar?n?n di?ilere oranla mimosine daha toleransl? olduklar? görülmü?tür (Santiago ve ark., 1988).
Kanavanin: Pek çok baklagil türünde bulunan bir termorezistants?z amino asit türüdür. Jack fasülyesi ve çay tohumu ile beslenen tilapyalarda, i?tah azalmas?, hareket yava?lamas? ve yüksek ölüm oran? gözlenmi?tir. Di?er bal?klar üzerindeki etkileriyle ilgili yeterli say?da çal??maya rastlan?lamam??t?r (Tacon, 1997).
Toksik Ya? Asitleri: Sterkulik ve malvalik asit gibi pamuk tohumu ya?? ve küspesinde bulunan siklopropan ya? asitlerinin, memelilerin üreme sistemlerinde anormalliklere ve ya? metabolizmas?nda de?i?imlere yol açt??? bilinmektedir. Bu maddelerin aflatoksin gibi di?er toksinlerle beraber, bal?klarda kanser yap?c? etkiye sebep olduklar? dü?ünülmektedir. Ayr?ca bu maddelerin gökku?a?? alabal?klar?nda, uzun zincirli ya? asidi metabolizmas?n? etkiledikleri ve stearik ve palmitik asit dehidrojenasyonunu engelledikleri görülmü?tür. Tam ya?l? pamuk tohumu küspesi içeren yemle beslenen tilapyalar?n, ya?s?z pamuk tohumu unu ile beslenenlere oranla daha az büyüdükleri görülmü?tür. Bu durum siklopropanoid ya? asitlerinin büyüme üzerindeki negatif etkilerine i?aret etmektedir (Robinson ve ark., 1984).
Mikotoksinler: Küfler taraf?ndan olu?turulan ikincil toksik bile?ikler olup yemlerde bulunan kimyasal etkenler içerisinde hayvan sa?l???n? tehdit eden en ciddi tehlikelerden biridir (Oruç, 2005). Yem hammaddeleri en önemli mikotoksin kayna??d?r. Yem hammaddeleri ve yemlerin depoland??? ko?ullara ba?l? olarak toksin kontaminasyonu, hammaddenin hasat?ndan ba?layarak, yemlerin bal?klara verilmesine kadar ki tüm a?amalarda meydana gelebilmektedir (Tekinay ve ark., 2005). Bal?k yeti?tiricili?inde mikotoksin kontaminasyon kayna?? m?s?r, f?nd?k unu, bu?day, soya ve pamuk küspesi gibi bitkisel kaynakl? hammaddelerdir. Mikotoksinler, bal?klar taraf?ndan biyolojik aç?dan önemli say?labilecek miktarlarda tüketilirlerse, canl? a??rl?k kayb?na, dü?ük döl verimine, kas yap?s?nda bozulmaya yol açt?klar? gibi ölüme bile neden olabilirler
Mikotoksinler, temizleme, y?kama, eleme, toksinle bula??k tohumlar?n veya danelerin ayr?lmas?, ?s? veya ???nlama gibi yöntemlerle yada kalsiyum hidroksit, sodyum bisülfit, monometilenamin, klorin gaz?, amonyak, hidrojen peroksit, amonyum hidroksit, hidroklorik asit ve formaldehit gibi baz? oksitleyici ve hidrolitik ajanlar kullan?larak yemden uzakla?t?r?labilmektedir.
Yukar?da ifade edilen bitkisel hammaddelerin içerdi?i antibesleyici faktörlerin, bal?klar üzerindeki etkilerini inceleyen çe?itli ara?t?rmac?lar?n elde ettikleri bulgular Tablo 1’de verilmi?tir.
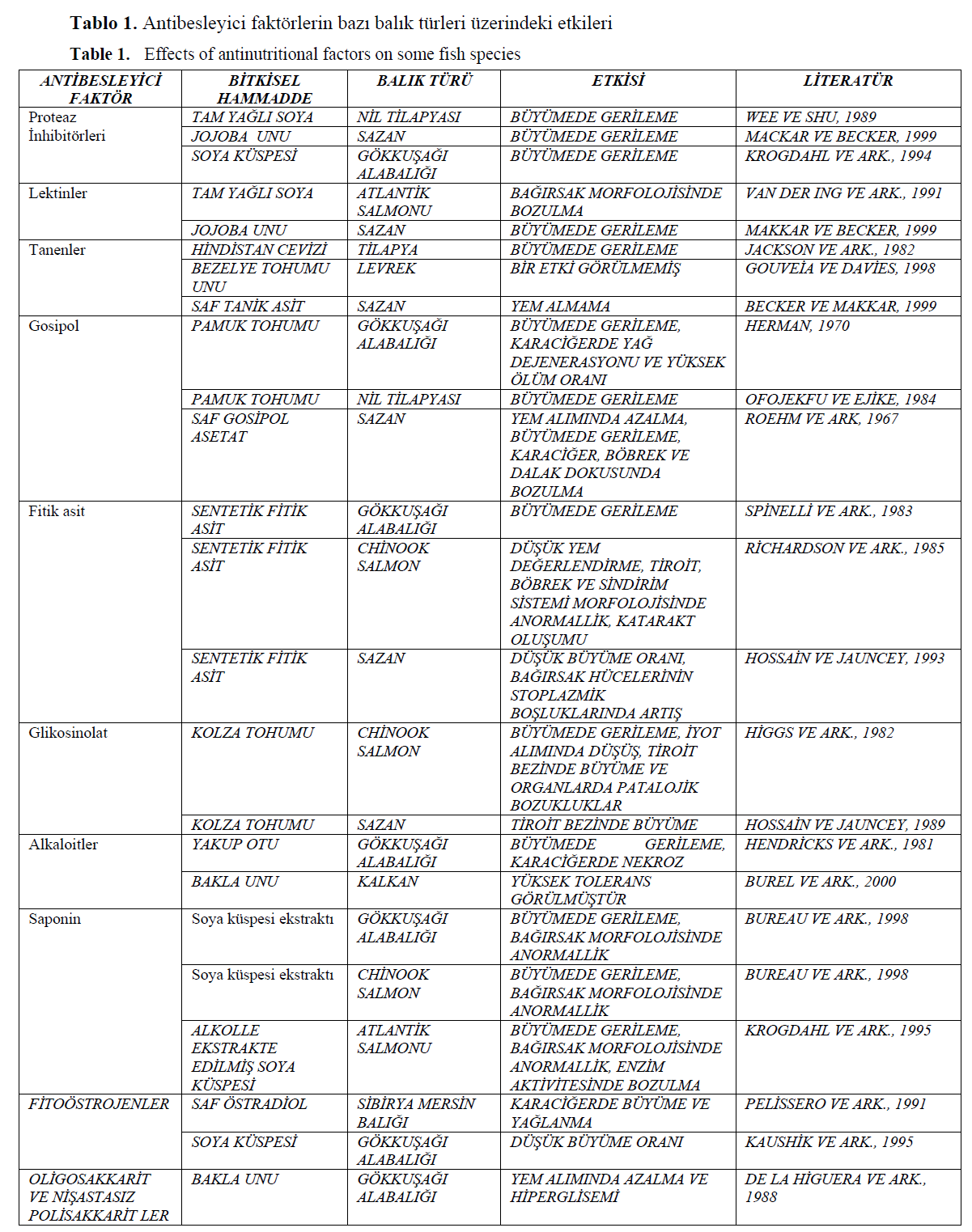
Tablo 1. Antibesleyici faktörlerin baz? bal?k türleri üzerindeki etkileri
Table 1. Effects of antinutritional factors on some fish species
Sonuç
Alternatif protein kaynaklar?n?n kullan?m? sonucu bal?k yemlerinde bulunan antibesleyici faktörler genelde ölüme yol açmasalar da, büyümede yava?lamaya ve üremede azalmaya sebep olabilmektedirler. Genel olarak proteaz inhibitörlerini, antijen maddeleri ve fitatlar? normal seviyede içeren yemler, bal?klar?n büyüme performans?n? etkilemezken, glukosinolatlar?, saponinleri, tanenleri, çözünebilir ni?astas?z polisakkaritleri ve gosipolleri içeren yemlerin ise yeti?tiricili?i yap?lan türlerin beslenmesinde daha önemli oldu?u görülmektedir. Lektin, fitoöstrojen ve alkaloit gibi maddeler ile ilgili ara?t?rmalar yeterli düzeyde olmad??? için, bu maddelerin, bal?klar?n fizyolojik ve ekolojik özellikleri üzerindeki etkilerini ortaya ç?karacak çal??malara ihtiyaç vard?r.
Bitkisel yemlerdeki antibesin oran?n? dü?ürmek için suyla ekstraksiyon, kuru ve özellikle de nemli ?s? yöntemleri, destek besinlerin eklenmesi gibi i?lemler ba?ar?l? bir ?ekilde uygulanmaktad?r. Yemin besin kalitesiyle ilgili negatif etkilerinden dolay? i?leme yöntemleri kullan?l?rken dikkatli olunmal?d?r. Örne?in, ?s? i?leminin protein ve karbonhidratlar?n kimyasal yap?lar?n? de?i?tirerek besin kalitesini dü?ürdü?ü bilinmektedir. Antibesin seviyelerinin dü?ürülmesi için i?lem uygulanmas?na karar verilmeden önce de?i?ik bal?k türlerinin antibesinlere olan farkl? tolerans seviyelerinin de dikkate al?nmas? gerekir. Örne?in, tilapya türlerinin genel olarak sazanlara oranla antibesin varl???na daha yüksek tolerans gösterdikleri söylenebilir. Safla?t?r?lm?? antibesinler tek ba??na kullan?larak yap?lan besleme deneyleri sayesinde kültür bal?klar?n?n üremelerini etkilemeyecek antibesin e?ik limitleri bulunabilir.
Çe?itli antibesleyici faktörler aras?ndaki etkile?im de dikkate al?nmas? gereken ba?ka bir önemli konudur. Baz? durumlarda bu etkile?im, antibesinlerin toksik etkilerinin azalmas?na yol açabilmektedir. Örne?in, saponin-tanen, tanenlektin ve tanen-siyanojen ikililerinin hepsinde etkile?ime giren antibesinlerin tek tek zehirli etkilerinin azald??? görülmü?tür. Bal?k yemi yap?m?nda birden fazla bitkisel hammaddenin kullan?ld??? dü?ünüldü?ünde bu hammaddeler içerisinde bulunan farkl? antibesleyici faktörlerin etkile?iminin nas?l sonuçland??? ve bal?klar üzerindeki etkisinin tan?mlanmas? için daha ayr?nt?l? ara?t?rmalara ihtiyaç duyulmaktad?r.
1160
References
- Akyıldız, R., (1992). BalıkYemleriveTeknolojisi. Ankara ÜniversitesiZiraatFakültesiYayınları: 1280, DersKitabı No: 366, Ankara. 192 s. Anonim, (2008). VikipediÖzgürAnsiklopedi. https://tr.wikipedia.org/wiki/Tanen (15.05.2008).
- Aregheore, E.M., Makkar, H.P.S., Becker, K., (1998). Assessment of lectin activity in a toxic and a non-toxic variety of Jatrophacurcas using latex agglutination and haemagglutination methods and inactivation of lectin by heat treatments, Journal of Science Food Agriculture, 77: 349–352.
- Balabanlı, C., Albayrak, S., Türk, M., Yüksel, O., (2006). Türkiyeçayırmeralarındabulunanbazızararlıbitkilervehayvanlarüzerineetkileri, SüleymanDemirelÜniversitesiOrmanFakültesiDergisi, 2: 89-96.
- Becker, K., Makkar, H.P.S., (1999). Effects of dietary tannic acid and quebracho tannin on growth performance and metabolic rates of common carp (Cyprinuscarpio L.), Aquaculture, 175: 327–335.
- Bilgiçli, N., (2002). Fitikasitinbeslenmeaçısındanönemivefitikasitmiktarıdüşürülmüşgıdaüretimmetotları, SelçukÜniversitesiZiraatFakültesiDergisi, 16 (30): 79-83
- Bureau, D.P., Harris, A.M., Cho, C.Y., (1998). The effects of purified alcohol extracts from soy products on feed intake and growth of chinook salmon (Oncorhynchustshawytscha) and rainbow trout (Oncorhynchusmykiss), Aquaculture, 161: 27–43.
- Burel, C., Boujard, T., Tulli, F., Kaushik, S.J., (2000). Digestibility of extruded peas, extruded lupin, and rapeseed meal in rainbow trout (Oncorhynchusmykiss) and turbot (Psetta maxima), Aquaculture, 188: 285–298.
- Büyüktuncer, Z., Başaran, A.A., (2005). Fitoöstrojenlervesağlıklıyaşamdakiönemleri, HacettepeÜniversitesi, EczacılıkFakültesiDergisi, 25 (2): 79-94.
- De la Higuera, M., Garcia-Gallego, M., Cardenete, G., Suarez, M.D., Moyano, F.J., (1988). Evaluation of Lupin seed meal as an alternative protein source in feeding ofrainbow trout (Salmogairdneri), Aquaculture, 71: 37–50.
- Ergün, A., Tuncer, Ş.D., Çolpan, İ. Yalçın, S., Yıldız, G., Küçükersan, M.K., Küçükersan, S., Önol, A.G., Muğlalı, Ö.H. veŞehu, A., (2002). Yemler, YemHijyeniveTeknolojisi. A.Ü. VeterinerFakültesi, HayvanBeslemeveBeslenmeHastalıklarıAnabilim Dalı, Ankara, 465 s
- . Ertaş, N., (2007). Yemeklikbaklagillerveantibesinselfaktörler, SelçukÜniversitesiZiraatFakültesiDergisi, 21 (41): 85-95.
- Gouveia, A., Davies, S.J., (1998). Preliminary nutritional evaluation of pea seed meal (Pisumsativum). For juvenile European sea bass (Dicentrarchuslabrax), Aquaculture, 166: 311–320.
- Francis, G., Makkar, P.S.H., Becker, K., (2001). Antinutritional factors present in plant derived alternate fish feed ingredients and their effects in fish, Aquaculture, 199: 197- 227.
- Hendricks, J.D., Sinnhuber, R.O., Henderson, M.C., Buhler, D.R., (1981). Liver and kidney pathology in rainbow trout Salmogairdneri exposed to dietary pyrrolizidine (Senecio) alkaloids. €xp.Mol.Pathol., 35:170-l83.
- In: Tacon, A., Basurco, B. (Eds)., Feeding Tomorrow’s Fish. Proceedings of the Workshop of the CIHEAM Network on Technology of Aquaculture in the Mediterranean (TECAM), Jointly Organised by CIHEAM, FAO and IEO, 24–26 June 1996, Mazarron, Spain. Cahiers-Options-Mediterraneennes, 22: 153–182.
- Herman, R.L., (1970). Effects of gossypol on rainbow trout Salmogairdneri Richardson, Journal of Fish Biology, 2: 293–303.
- Higgs, D.A., McBride, J.R., Markert, J.R., Dosanjh, B.S., Plotnikoff, D.M., Clarke, C.W., (1982). Evaluation of Tower and Candle rapeseed (canola) meal and Bronowski rapeseed protein concentrate as protein supplements in practical dry diets for juvenile chinook salmon (Oncorhynchustshawytscha), Aquaculture, 29: 1–31.
- Hossain, M.A., Jauncey, K., (1989). Studies on the protein, energy and aminoacid digestibility of fishmeal, mustard oilcake, linseed and sesame meal for common carp(Cyprinuscarpio L.), Aquaculture, 83: 59– 72.
- Hunt, A.Ö., Özkan, F., Altun, T., (2007). Balıkyemlerindebeslenmeyisınırlandırıcımaddelerveetkileri, TürkSuculYaşamDergisiUlusal Su Günleri 2007Özel Sayı, 5-8: 634- 642.
- Jackson, A.J., Capper, B.S., Matty, A.J., (1982). Evaluation of some plant proteins in complete diets for the tilapia (Sarotherodonmossambicus). Aquaculture, 27: 97–109.
- Kaya, İ., Yalçın, S., (1999). Baklagiltaneyemlerive ruminant rasyonlarındakullanımı, LalahanHayvanaraştırmaEnstitüsüDergisi, 39: 101-114.
- Kaushik, S.J., Cravedi, J.P., Lalles, J.P., Sumpter, J., Fauconneau, B., Laroche, M., (1995). Partial or total replacement of fishmeal by soybean protein on growth, protein utilization, potential estrogenic or antigenic effects, cholesterolemia and flesh quality in rainbow trout, Oncorhynchusmykiss, Aquaculture, 133: 257–274.
- Krogdahl, A., Lea, T.B., Olli, J.J., (1994). Soybean proteinase inhibitors affect intestinal trypsin activities and amino acid digestibilities in rainbow trout (Oncorhynchusmykiss), Comparative Biochemistry and Physiology, 107A: 215– 219.
- Krogdahl, A., Bakke-Mckellep, A.M., Roed, K.H., Baeverfjord, G., (2000). Feeding Atlantic salmon Salmosalar L. soybean products: effects on disease resistance (furunculosis), and lysozyme and IgM levels in the intestinal mucosa, Aquaculture Nutrition, 6: 77–84.
- Makkar, H.P.S., Becker, K., (1999). Nutritional studies on rats and fish (carp, Cyprinuscarpio). fed diets containing unheated and heated Jatrophacurcas meal of a non-toxic provenance, Plant Foods for Human Nutrition, 53: 183–192.
- Midilli, M., Muğlalı, H., Alp, M, Kocabağlı, N., Tanör, M.A., Toklu, G.S., (2003). Yemekatılanfitazenzimininbroylerlerdebesiperformansıve mineral dengesiüzerineetkisi, Turkish Journal of Veterinary and Animal Science, 27: 751-759.
- Ofojekwu, P.C., Ejike, C., (1984). Growth response and feed utilisation in the tropical cichlid Oreochromisniloticusniloticus (Linn). fed on cottonseed-based artificial diets, Aquaculture, 42: 27–36.
- Oruç, H. H., (2005). MikotoksinlerveTanıYöntemleri. Uludag University. Journal of The Faculty of Veterinary Medicine, 24: 1-2- 3-4: 105-110.
- Pekşen, E. veArtık, C., (2005). Antibesinselmaddelerveyemekliktanebaklagillerinbesleyicideğerleri, OMÜ ZiraatFakültesiDergisi, 20 (2) : 110-120.
- Pelissero, C., Le Menn, F., Kaushik, S., (1991). Estrogenic effect of dietary soybean meal on vitellogenesis in cultured Siberian sturgeon Acipenserbaeri, Gen. Comp. Endocrinology, 83: 447–457.
- Refstie, S., Svihus, B., Shearer, K.D., Storebakken, T., (1999). Nutrient digestibility in Atlantic salmon and broiler chickens related to vscosity and non-starch polysaccharide content in different soybean products, Animal Feed Science Technology, 79: 331–345.
- Richardson, N.L., Higgs, D.A., Beames, R.M., McBride, J.R., (1985). Influence of dietary calcium, phosphorous, zinc and sodium phytate level on cataract incidence, growth and histopathology in juvenile chinook salmon (Oncorhynchustshawytscha).
- In: Francis, G., Makkar, Harinder P.S., Becker, K., 2007. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fis,. Aquaculture 199, 197–227.
- Robinson, E.H., Rawles, S.D., Oldenburg, P.W., Stickney, R., (1984). Effects of feeding glandless or glanded cottonseed products and gossypol to Tilapia aurea, Aquaculture, 38: 145–154.
- Roehm, J.N., Lee, D.J., Sinnhuber, R.O., (1967). Accumulation and elimination of dietary gossypol in the organs of rainbow trout, Journal of Nutrition, 92: 425–428.
- Rumsey, G.L., Hughes, S.G., Winfree, R.A., (1993). Chemical and nutritional evaluation of soy protein preparations as primary nitrogen sources of rainbow trout (Oncorhynchusmykiss). Animal Feed Science Technology, 40: 135–151.
- Santiago, C.B., Aldaba, M.B., Laron, M.A., Reyes, O.S., (1988). Reproductive performance and growth of Nile tilapia (Oreochromisniloticus) broodstock fed diets containing Leucaenaleucocephala leaf meal, Aquaculture, 70: 53–61.
- Seyrek, K., Bildik, A., (2001). Lektinler, YüzüncüYılÜniversitesiVeterinerlikFakültesiDergisi, 12: 96-100
- Spinelli, J., Houle, C.R., Wekell, J.C., (1983). The effect of phytates on the growth of rainbow trout (Salmogairdneri). fed purified diets containing varying quantities of calcium and magnesium, Aquaculture, 30: 71-83.
- Storebakken, T., Shearer, K.D., Roem, A.J., (1998). Availability of protein, phosphorus and other elements in fishmeal, soy protein concentrate and phytase-treated soy proteinconcentrate- based diets to Atlantic salmon, Salmosalar, Aquaculture, 161: 365–379.
- Tacon, A.G.J., (1997). Fishmeal replacers: review of antinutrients within oilseeds and pulses—a limiting factor for the aquafeed Green Revolution? In: Tacon, A., Basurco, B. (Eds).,
- Feeding Tomorrow’s Fish. Proceedings of the Workshop of the CIHEAM Network on Technology of Aquaculture in the Mediterranean (TECAM), Jointly Organised by CIHEAM, FAO and IEO, 24–26 June 1996, Mazarron, Spain. Cahiers-Options-Mediterraneennes, 22: 153–182.
- Tekinay, A.A., Güroy, D., Çevik, N., (2005). Balıkyemlerindemikotoksinkontrolü. (Bildiriözeti), TürkSuculYaşamDergisiUlusal Su Günleri 2005 SempozyumuÖzelSayısı, 4: 538.
- Van der Ingh, T.S.G.A.M., Krogdahl, A., Olli, J.J., Hendriks, H.G.C.J.M., Koninkx, J.F.J.G., (1991). Effects of soybeancontaining diets on the proximal and distal intestine in Atlantic salmon Salmosalar : a morphological study, Aquaculture, 94: 297– 305.
- Wee, K.L., Shu, S.-W., (1989). The nutritive value of boiled full-fat soybean in pelleted feed for Nile tilapia, Aquaculture, 81: 303– 314.


