Ying Chen1,2, Hui Gao1,2 and Hui Zhao1*
1Department of Chemistry and Environmental Science, Guangdong Ocean University, Zhanjiang, China
2Department of Southern Marine Science and Engineering Guangdong Laboratory, Zhuhai, China
3Department of Marine Sciences, Nanjing University of Information Science and Technology, Nanjing, China
- *Corresponding Author:
- Hui Zhao
Department of Chemistry and Environmental Science, Guangdong Ocean University, Zhanjiang, China
E-mail: huizhao1978@163.com
Received Date: 15.02.2021; Accepted Date: 02.03.2021; Published Date:09.03.2021
Phytoplankton as a key element in the marine food chain plays a vital role in marine ecosystems. In the South China Sea, phytoplankton blooms often occur northwest of Luzon Island usually during winter, which may have an important impact on primary productivity and regional biological carbon pump. Located in the East Asian monsoon region, the region northwest of Luzon Island is affected by wind stress, Kuroshio and topography with complex dynamic environment. With comprehensive analysis of domestic and international recent studies on phytoplankton blooms in the region northwest of Luzon Island, it is in order to explore the significant impacts of the environmental conditions including wind, Cagayan river discharge, Kuroshio intrusion, upwelling and currents on the spatio-temporal distribution of phytoplankton blooms. It is concluded that the winter phytoplankton bloom is likely due to upwelling and turbulent mixing generated by the Kuroshio interaction with the topography of the Luzon Island. The Cagayan river discharge also contributes to this occurrence, which is the largest river in the Philippines. But the impact of the river is limited to a few kilometers from its mouth. Moreover, the northward flow west of Luzon Island causes the center of the phytoplankton bloom to move northward. This review outlines the impacts of environmental elements on winter phytoplankton blooms northwest of the Luzon Island, and explores potential formation mechanisms of the blooms. Its spatio-temperal distribution can reveal the changes of marine ecosystem indirectly and is of great significance to marine scientific research. The researches on winter phytoplankton blooms also play a guiding role in red tide disaster prevention, marine environmental protection research, fisheries development, to some degree. The more detailed knowledge of the dynamic mechanisms of phytoplankton bloom phenomena contributes to better understanding of the complex marine biogeochemical process near the Luzon Strait.
Keywords
Winter phytoplankton blooms; Upwelling; Ekman transport; Kuroshio intrusion; Luzon Island
Introduction
Marine phytoplankton is the basis of the marine food chains and affects sea-surface carbon dioxide flux through photosynthesis, which contributes roughly half of the biospheres net primary production [1,2]. Marine primary productivity plays an important role in the exchange of carbon dioxide at the air-sea interface, global carbon cycle and global climate change. Marine phytoplankton biomass, commonly quantified as the concentration of the phytoplankton pigment chlorophyll-a (Chl-a), is an important indicator of eutrophication in marine ecosystems [3]. Chl-a is also an important parameter for evaluating marine water quality and for detecting marine fisheries. With basic ecological information, its spatiotemporal variation is closely related to many environmental factors, such as temperature, light conditions, photosynthetically active radiation, nutrients, wind field and current field [4-7]. Spatiotemporal variations of phytoplankton has a profound impact on marine fisheries and global biogeochemical cycles [8-10] and understand the phytoplankton variation may help us to better comprehend the basic ecological environments and their responses to global climate change [11-13].
The South China Sea (SCS), with a total area of about 3.5 × 106 km2, is a large semi-enclosed tropical marginal sea in the western Pacific Ocean [14]. The Luzon Strait has a width of about 250 km and has a sill depth of about 2400 m [15]. It is the main channel between the western Pacific Ocean and the SCS, whereas other channels are either too shallow or too narrow for effective water transport [16,17]. The hydrological characteristics of Luzon Strait are complex and influenced by the general condition of the SCS, which are in turn influenced by the Asian monsoon system [18]. Mesoscale eddies occur frequently in the SCS [19], while intense activities of eddies appeared also in the region northwest of Luzon [20,21]. Upwelling caused by the cyclonic eddies has significant effects on the vertical transport of nutrients and distribution of phytoplankton in the upper ocean. The upwelling regions account for only 1% of the whole ocean, but influence of upwelling on the ocean is far-reaching. It plays an important role in the redistribution of marine nutrients. Chavez et al. [22] estimated that 67% of global new productivity was provided by the upwelling system, about 4.8 Gt C yr-1. The mean SCS primary productivity is only about 350 mg m−2d−1 [4], but in the vicinity of the Luzon Strait primary productivity is over 600 mg m−2 d−1 [23], with high surface Chl-a in winter[18,23-26].
In recent years, there were a number of case studies on the phytoplankton blooms northwest of Luzon Island. Most of the studies examined roles of wind stress curls, mixedlayer entrainments, coastal currents, Kuroshio intrusion in individual winter bloom and interannual variability of the winter blooms [2,18,23-29]. The review outlines influence of environmental elements on the winter phytoplankton blooms northwest of Luzon Island. By analyzing the existing research results, this review also discusses the potential formation mechanism of phytoplankton blooms northwest of the Luzon Island, which may help better understanding regional primary productivity of marine phytoplankton systematically, providing a reference for further researches on primary productivity of phytoplankton in this region.
Possible Environmental Factors Regulating the Winter Phytoplankton Blooms
The influence of wind
The SCS is predominantly controlled by the East Asian monsoon, with strong northeasterly winds prevailing from November to March [4]. Wind events may play an important role in physical and biological processes of the upper ocean. Historical researches showed that the average wind speed is approximately 9 ms-1 (~0.26 Pa) during the winter in the SCS [30], and the highest wind speed exceeds 10 ms-1 northwest of Luzon Island [29]. Chen [23] confirmed the high Chl-a appears northwest of Luzon Island in winter using in situ measured data. The results showed that the average Chl-a near the Luzon Strait under the northeasterly monsoonal winds in winter is as high as eight times of the average Chl-a concentration in summer. It is worth noting that there are also more mineral dusts and atmospheric nitrogen depositions into the SCS transported by the northeasterly wind during winter [31,32]. The research indicated that the surface mixed layer depth northwest of Luzon Island deepened from ~12 m in summer to more than 60 m in winter [18]. The strong wind during the northeasterly monsoonal wind would generally enhance stratification of upper ocean due to sea surface cooling, intensify mixing, and increase entrainment, and trigger high Chl-a concentration. The wind-driven Ekman pumping as well as entrainment mixing would also bring nutrients upward to the euphotic zone.
The influence of Kuroshio intrusion
The Kuroshio intrudes from the Philippine Sea into the SCS through the Luzon Strait, which is an area where strong vertical mixing occurs due to the Kuroshio intrusion as well as strong wind. A cyclonic mesoscale eddy is formed in the northeast of the SCS under the winter monsoon and sea floor topography. It leads to the shallowness of the nutricline [4,33] to 50-70 m [34] and results in higher phytoplankton productivity [33,35]. For example, the ratio of nitrateuptake- based new production to primary production is approximately 50% in winter and 30% in summer within the euphotic zone of the Kuroshio water [36]. The Kuroshio can uplift the subsurface high-nutrient water to the surface and stimulate phytoplankton growth, which is hypothesized by Peñaflor [25] and is evidenced in Guo et al. [28] study. The winter phytoplankton blooms form frequently to the west of the Luzon Strait [4,23-25]. The blooms are characterized by an inverted-v shape fronts between the Kuroshio and SCS water and the Luzon coastal current [25,28]. The blooms are clearly separated from relatively high Chl-a in the SCS by the Kuroshio. Previous studies indicated that winter phytoplankton blooms were found near the Luzon Strait, and the phytoplankton blooms in this area were related to the mesoscale eddies caused by the Kuroshio intrusion [26,28,35] showed that stronger Kuroshio intrusion resulted in stronger fronts, and then intensified upwelling and enhanced Luzon winter blooms during El Niño years, which provided new insights into the impact of Kuroshio on the SCS ecosystem. Moreover, the Kuroshio intrusion and northward current along the west Luzon also play important role in nutrient horizontal transport. Kuroshio intrusion into the SCS occurs almost all year round, while phytoplankton blooms rarely occur in all seasons other than winter. It may be due to that the winter mixing deepens the depth of mixing layer, which provides a favorable condition of nutrient uplift for the nutrient availability of the phytoplankton blooms.
The influence of upwelling and northward flow
The upwelling promotes the mixed exchange of deep water and surface water. Xue [37] showed that the Ekman transport-induced movement of Kuroshio water during winter monsoon generated upwelling. Zhao et al. [2] observed a strong upwelling tendency from Ekman pumping velocity (>1 × 10−5 ms−1) in the northwest of Luzon, and conjectured that wind-induced upwelling (Ekman pumping) and entrainment mixing may be important factors leading to the high winter Chl-a. Wang et al. [18] also demonstrated the winter phytoplankton bloom induced by both Ekman pumping-driven upwelling and upper mixed layer entrainment in the southwest of Luzon Strait. Previous studies indicated that the upwelling was responsible for the winter phytoplankton blooms southwest of Luzon Strait. Winter upwelling brings nutrients from deeper layers to the surface, thereby increasing phytoplankton reproduction [2,18,23-25,27,35,38]. A novel finding in Gao et al. [29] is that the advection effect of the northward background coastal flow induces the inconsistency between the upwelling center and winter bloom center. There exists a strong northward coastal current west of Luzon in winter [39,40]. The northward movement of northward coastal current west of Luzon Island has a certain effect on the difference between the wind-induced upwelling and locations of phytoplankton blooms. The upwelling brings the abundant nutrients at the bottom to the surface layer during the winter monsoon, leading to the winter phytoplankton blooms. In addition, the surface flow carries phytoplankton northward, causing the phytoplankton bloom center to move northward from the upwelling center induced by Ekman transport.
The influence of Cagayan river discharge
Although upwelling is the main factor of phytoplankton blooms as mentioned above, it is still necessary to investigate the possible impact of Cagayan river discharge into the region. Another factor that may influence the phytoplankton blooms formation in the region northwest of Luzon Island is the nutrients coming from the Cagayan river. The Cagayan river is the largest river in the Philippines and its flow ranges between 180 m3/s in August and 500 m3/s in December and the period of highflow is from October to December [26]. On some occasions, nearshore blooms are substantially enhanced by nutrient input from the Cagayan river. Penaflor et al. [25] firstly indicated that the Chl-a levels were constantly high throughout the year with the potential influences of Cagayan river discharge in the vicinity of the northern Luzon Island coast. Results also present that the Cagayan river influence is only limited near the northern coast of Luzon Island within about few kilometers from the river mouth under the influence of a Kuroshio branch. Most previous researches have been ignored this phenomenon. In addition, under the influence of the northeast monsoon, the coastal water produced by Cagayan river discharge on the north coast of Luzon Island may have been advected into the phytoplankton blooms region. The blooming season mostly coincides with the flooding season of the Cagayan river [26]. The fresh water from Cagayan river discharge would carries a large amount of terrestrial nutrients, resulting in higher Chl-a and higher primary productivity. In addition, weather events such as rainfall would increase the nutrients input from the Cagayan river in the region north of Luzon Island.
Mechanisms Driving the Winter Phytoplankton Blooms
The schematic diagrams presented in Figure 1 illustrate the mechanisms underlying the offshore winter phytoplankton bloom in the region northwest of Luzon Island. On one hand, strong winter monsoon facilitates evaporative cooling to enhance mixing in the upper ocean. On the other hand, the tendency of turbulence to diffuse into the adjoining non-turbulent layer leads to the destruction of the underlying thermocline stratification, and increases the thickness of the mixed layer by enhancing the entrainment [41-43]. The intensified mixing and increased entrainment induce subsurface upwelling, which brings nutrients from the deep sea and support the growth of phytoplankton from the subsurface upwelled water to the upper layer [44-46]. The phytoplankton blooms are as well as substantially enhanced by nutrient input from the Cagayan river in the region north of Luzon Island. In addition, weather events such as rainfall would increase the nutrients input from the Cagayan river, increasing the intensity of phytoplankton blooms. The Kuroshio also plays a major role in the formation of the winter phytoplankton Blooms. Because of the mesoscale processes generated by the Kuroshio intrusion, and the interaction between Kuroshio and topography, the bloom is mainly triggered and sustained by nutrient pumping and entrainment from deep depth in winter. Affected by the advection effect of the geostrophic flow, the center of the phytoplankton bloom area shifts northward.
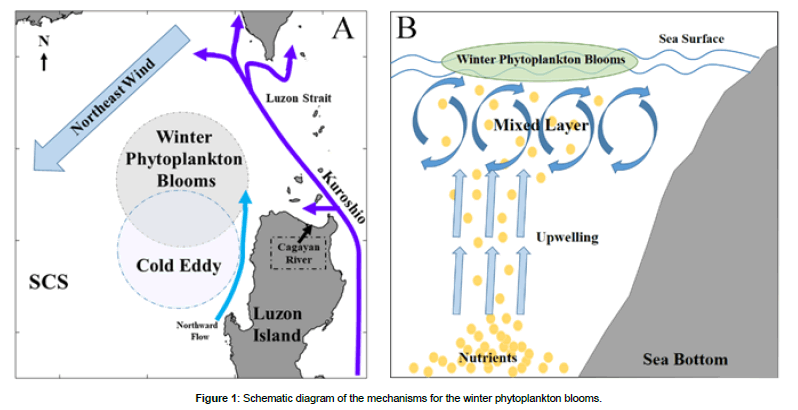
Figure 1: Schematic diagram of the mechanisms for the winter phytoplankton blooms.
Discussion and Conclusion
This review mainly describes the impact of environmental elements on the winter phytoplankton blooms northwest of the Luzon Island and the potential formation mechanism of the blooms. In summary, we attribute the blooms based on previous studies to the winter wind, as well as upwelling and turbulent mixing, which are generated by the Kuroshio interaction and the Kuroshio-topography interaction. Nutrient input from the Cagayan river may also affect phytoplankton blooms in the north of Luzon Island, and rainfall may deepen this phenomenon. The advection effect of the northward background coastal flow induces the inconsistency between the upwelling center and winter bloom center, and the blooms center would move northward. However, previous studies mainly focus on the sea surface, and dynamic process of the marine threedimensional structure needs to be further investigated. Due to the limitation of in situ measured data, the ocean numerical model is a good research tool. Future researches should pay more attention to the combination of numerical models and observations to analyze the blooms caused by marine mesoscale eddies and sub-mesoscale phenomena with different three-dimensional structures near Luzon Island.
Author’s Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approve it for publication.
Funding
The present research is supported by National Natural Science Foundation of China (No. 42076162) and the Natural Science Foundation of Guangdong Province, China (Grant No. 2020A1515010496).
36175
References
- Behrenfeld MJ, Malley RT, Siegel DA, McClain CR, Sarmiento JL, et al. (2006) Climate-driven trends in contemporary ocean productivity. J Nature 444: 752-755.
- Zhao H, Sui D, Xie Q, Han G, Wang D, et al. (2012) Distribution and interannual variation of winter phytoplankton blooms northwest of Luzon Islands from satellite observations. J Aquat Ecosyst Health Manag 15: 53-61.
- Smith VH (2006) Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnol Oceanogr 51: 377-384.
- Liu KK, Chao SY, Shaw PT, Gong GC, Chen CC, et al. (2002) Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study. J Deep Sea Res Part I Oceanog Res. 49: 1387–1412
- Zhao, H., Tang, D. L. (2007) Effect of 1998 El Niño on the distribution of phytoplankton in the South China Sea. J Geophys Res 112: 117-128.
- Lin LR, Zhao H (2012) Analysis on the relations between sea surface temperature and phytoplankton Chlorophyll-a in the South China Sea. J Mar Sci 30: 46-54.
- Zhao H, Tang DL, Wang YQ (2008) Comparison of phytoplankton blooms triggered by two typhoons with different intensities and translation speeds in the South China Sea. J Mar Ecol Prog Ser 365: 57-65.
- Falkowski PG (1998) Biogeochemical controls and feedbacks on ocean primary production. J Science 281:200-206.
- Falkowski PG, Oliver MJ (2007) Mix and match: How climate selects phytoplankton. Nat Rev Microbiol 5: 813-819.
- Chassot E, Bonhommeau S, Dulvy NK, Melin F, Waston R, et al. (2010) Global marine primary production constrains fisheries catches. J Ecology Letters 13: 495-505.
- Thomas AC, Mendelssohn R, Weatherbee R (2013) Background trends in California Current surface chlorophyll concentrations: A state-space view. J Geophy Resear: Oceans 118: 5296-5311.
- Boyce DG, Lewis MR, Worm B (2010) Global phytoplankton decline over the past century. J Nature 466: 591-596.
- Antoine D, Morel A, Gordon HR, Banzon VF, Evans RH (2005) Bridging ocean color observations of the 1980s and 2000s in search of long-term trends. J Geophy Res Oceans 110.
- Zhao H, Wang Y (2018) Phytoplankton increases induced by tropical cyclones in the South China Sea during 1998–2015. J Geophys Res 123: 2903-2920.
- Zhou C, Zhao W, Tian J, Yang Q, Qu T (2014) Variability of the deep-water overflow in the luzon strait. J Phys Oceanogr 44: 2972–2986.
- Chen YL, Chen HY, Karl DM,Takahashi M (2004) Nitrogen modulates phytoplankton in spring in the South China Sea. J Continental Shelf Research 24: 7-541.
- Metzger EJ, Hurlburt HE (2001) The non-deterministic nature of the Kuroshio penetration and the eddyshedding in the South China Sea. J Phys Oceanngrp 31: 1712-1732.
- Wang JJ, Tang DL, Sui Y (2010) Winter phytoplankton bloom induced by subsurface upwelling and mixed layer entrainment southwest of Luzon Strait. J Mar Syst 83: 141-149.
- Zheng QA, Xie LL, Zheng ZW, Hu JY (2017) Progress in Research of Mesoscale Eddies in the South China Sea. J Adv Mar Sci 35: 131-158.
- Chen G, Hou Y, Chu X (2011) Mesoscale eddies in the South China Sea: Mean properties, spatiotemporal variability and impact on thermohaline structure. J Geophys Res Oceans 116: 1-19.
- Wang G, Su J, Chu PC (2003) Mesoscale eddies in the South China Sea observed with altimeter data. J Geophys Resear Letters 30: 1-6.
- Chavez FP, Messié M, Pennington JT (2011) Marine primary production in relation to climate variability and change. J Ann Rev Mar Sci 3: 227
- Chen CC, Shiah FK, Chung SW, Liu KK (2006) Winter phytoplankton blooms in the shallow mixed layer of the South China Sea enhanced by upwelling. J Mar Syst 59: 97-110
- Tang DL, Ni IH, Kester DR, Karger MFE (1999) Remote sensing observations of winter phytoplankton blooms southwest of the luzon strait in the South China Sea. J Mar Ecol Prog 191: 43-451.
- Peñaflor EL, Villanoy CL, Liu CT, David LT (2007) Detection of monsoonal phytoplankton blooms in Luzon Strait with MODIS data. J Remote Sens Environ 109: 443-450.
- Shang S, Li L, Li J, Li Y, Lin G, et al. (2012) Phytoplankton bloom during the northeast monsoon in the Luzon Strait bordering the Kuroshio. J Remote Sens Environ 124: 38-48.
- Lu W, Yan XH, Jiang Y (2015) Winter bloom and associated upwelling northwest of the Luzon Island: A coupled physical-biological modeling approach. J Geophys Res Oceans 120: 533-546.
- Guo L, Xiu P, Chai F, Xue HJ, Wang DX, et al. (2017) Enhanced chlorophyll concentrations induced by Kuroshio intrusion fronts in the northern South China Sea. J Geophys Res Lett 44: 11565-11572.
- Gao H, Zhao H, Han G, Dongm C (2021) Spatio-Temporal variations of winter Phytoplankton blooms Northwest of the Luzon Island in the South China Sea. Front Mar Sci 8: 637499
- Wyrtki K (1961) Physical oceanography of the Southeast Asian waters NAGA Rep 2: 195 Scripps Inst Oceanogr Univ Calif San Diego.
- Fanning KA (1989) Influence of atmospheric pollution on nutrient limitation in the ocean. J Nature 339: 460-463.
- Merrill JT, Uematsu M, Bleck R (1989) Meteorological analysis of long range transport of mineral aerosols over the North Pacific. J Geophys Res 94: 8584-8598.
- Shaw PT, Chao SY (1994) Surface circulation in the South China Sea. J Deep Sea Research Part I: Oceanogr Resear Papers 41: 1663-1683.
- Tseng CM, Wong GTF, Lin II, Wu CR, Liu KK (2005) A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea. J Geophy Resear Letters 32: 487-500.
- Chen LYL, Chen HY, Lin II, Lee MA, Chang J (2007) Effects of cold eddy on phytoplankton production and assemblages in Luzon Strait bordering the South China Sea. J Oceanogr 63: 671-683
- Chen YL, Chen HY, Tuo SH, Ohki K (2008) Seasonal dynamics of new production from Trichodesmium N2 fixation and nitrate uptake in the upstream Kuroshio and South China Sea basin. Limnol Oceanogr 53: 1705-1721.
- Xue H, Chai F, Pettigrew N, Xu D, Shi M, et al. (2004) Kuroshio intrusion and the circulation in the South China Sea. J Geophys Resear 109 C02017.
- Qiu C, Wang D, He Z, Cheng C (2012) Seasonal variability of chlorophyll a fronts in the Luzon Strait based on satellite observations. J Aquatic Eco Health Manag15: 46-52.
- Chao SY, Shaw PT, Wu SY (1996) Deep water ventilation in the South China Sea. J Deep Sea Res Part Oceanogr Res Pap 43: 445–466.
- Shaw PT, Chao SY, Liu KK, Pai SC, Liu CT (1996) Winter upwelling off Luzon in the northeastern South China Sea. J Geophys Res Oceans 101: 16435-16448.
- Strang EJ, Fernando HJS (2001) Entrainment and mixing in stratified shear flows. J Fluid Mech 428: 349-386
- Backhaus JO, Hegseth EN, Wehde H, Irigoien X, Hatten K, et al. (2003) Convection and primary production in winter. J Ma Ecol Prog Ser 251: 1-14.
- Labiosal RG, Arrigo KR (2003) The interplay between upwelling and deep convective mixing in determining the seasonal phytoplankton dynamics in the Gulf of Aqaba: Evidence from SeaWiFS and MODIS. J Limnol Oceanogr 48: 2355-23368.
- Madhupratap M, Kumar SP, Bhattathiri PMA, Kumar MD, Raghukumar S, et al. (1996) Mechanism of the biological response to winte rcooling in the northeastern Arabian Sea. J. Nature 384: 549–552.
- Li L, Wu R, Guo X (2000) Seasonal circulation in the South China Sea-a TOPEX/POSEIDON satellite altimetry study. J Acta Oceanologica Sinica 22.
- Zhao H, Han G, Zhang S, Wang DX (2013) Two phytoplankton blooms near Luzon Strait generated by lingering Typhoon Parma. J Geophy Resear: Biogeosci 118: 412-421.


